Воспроизведение клетки
Клеточный цикл. Все новые клетки возникают в результате деления надвое уже существующих. Многоклеточный организм также начинает свое развитие чаще всего с одной-единственной клетки. Путем многократных делений образуется огромное количество клеток, которые и составляют организм.
В многоклеточных организмах не все клетки способны к делению по причине их высокой специализации. Именно такими являются клетки, имеющие разную продолжительность жизни. Например, нервные и мышечные клетки после завершения эмбрионального периода развития организма перестают делиться и функционируют на протяжении всей последующей жизни организма. Многие специализированные клетки не размножаются в обычных условиях, но при повреждении органов и тканей, в состав которых они входят, их способность к делению восстанавливается. К таким клеткам относятся клетки печени, эндокринных желез и других органов. Другие клетки, в частности костного мозга, эпителия тонкого кишечника, эпидермиса, в процессе выполнения своей специфической функции погибают, поэтому на смену им приходят новые, образованные путем деления.
Постоянное деление клеток верхушечной и боковых меристем растений (камбия, пробкового камбия) обеспечивает длительный рост побегов и корней в длину и толщину.
Вновь образованные клетки приобретают способность к делению после некоторого периода роста. Кроме того, делению предшествует удвоение клеточных структур: хлоропластов, митохондрий, центриолей и др.
Перед началом клеточного деления должна реплицироваться ДНК, поскольку она несет в себе информацию, необходимую клетке для синтеза белков. Если бы дочерние клетки не получали точных копий ДНК материнской клетки, они перестали бы быть похожими друг на друга. Это в конечном итоге привело бы к невозможности существования определенных видов организмов. Чтобы этого не случилось, ДНК должна идеально реплицироваться и каждая дочерняя клетка при клеточном делении должна получать ее копию.
Совокупность последовательных и взаимосвязанных процессов в период подготовки клетки к делению и в период деления называется митотическим циклом (от названия основного типа деления —митоза).
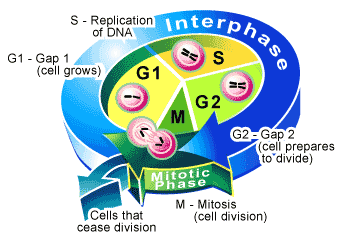
Жизнь клетки от одного деления до следующего или до смерти называется клеточным или жизненным циклом клетки. У одноклеточных организмов клеточный цикл совпадает с жизнью особи. В непрерывно размножающихся тканевых клетках клеточный цикл совпадает с митотическим циклом и состоит из четырех периодов со строгой последовательностью смен друг друга: постмитотического (G1 — англ. grow — расти, увеличиваться), синтетического (S — англ. synthesis — синтез), премитотического(G2) амитоза (М) (рис. 1.23). Первые три периода —это интерфаза. По продолжительности она составляет большую часть митотического цикла клетки.
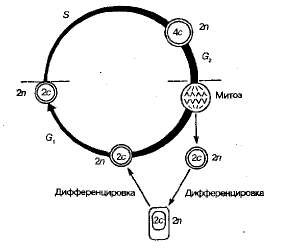
Рис. 1.23. Схема митотического цикла.
В периоде G, клетка растет, в ней усиленно образуются РНК и белки, в первую очередь ферменты, катализирующие образование предшественников ДНК, ферменты метаболизма РНК и строительных белков.
В 5-периоде происходит репликация ДНК (синтез новых цепочек ДНК — отсюда и название периода). В результате удвоения молекул ДНК в каждой хромосоме оказывается вдвое больше ДНК, чем было до S-периода, т. е. количество ДНК в диплоидных клетках соответствует тетраплоидному набору. Однако количество хромосом считается диплоидным, потому что образовавшиеся дочерние хромосомы (хроматиды) тесно переплетены между собой и составляют единую хромосому.
В S-периоде, кроме репликации ДНК, продолжается синтез РНК и белков, осуществляется синтез рРНК, которые используются в следующем периоде для синтеза белков, обеспечивающих митоз, происходит удвоение центриолей, а также деление митохондрий, увеличивается количество других органелл клетки.
Время от окончания синтеза ДНК и до начала митоза называется G2-периодом. В этот период завершается подготовка клетки к митозу, активно синтезируются различные белки, в том числе и тубулиновые, из которых образуются нити веретена деления, а также иРНК и белки для осуществления (G1-периода после митоза. Далее наступает деление клетки.
Деление клетки. После того как в клетке завершаются биохимические процессы подготовки к делению, начинается таинственный и до конца не изученный процесс. В настоящее время известно несколько способов деления клетки: митоз, прямое бинарное деление, амитоз и мейоз.
Митоз (греч. mitos — нить), или непрямое деление, — основной способ деления эукариотических клеток. Митоз — это деление клетки, приводящее к образованию двух дочерних, в каждой из которых имеется точно такой же (аналогичный) набор хромосом, как и в родительской. Митоз — процесс непрерывный, но для удобства изучения биологи делят его на четыре стадии в зависимости от того, как выглядят в это время хромосомы в световом микроскопе. В митозе выделяют профазу, метафазу, анафазу и телофазу (рис. 1.24).
В профазе происходит укорочение и утолщение хромосом вследствие их спирализации. В это время хромосомы двойные; они состоят из двух хроматид, связанных между собой в области первичной перетяжки кинетохором. Удвоение хромосом, как говорилось, произошло в 5-периоде интерфазы. Одновременно с утолщением хромосом исчезает ядрышко и фрагментируется (распадается на отдельные цистерны) ядерная оболочка. После этого хромосомы лежат в цитоплазме свободно и беспорядочно.
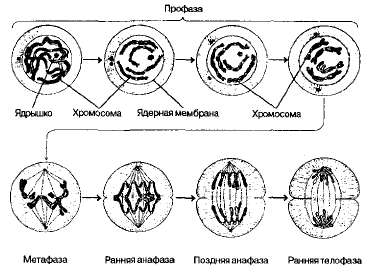
Рис. 1.24. Схема митоза.
В профазе центриоли (в тех клетках, где они есть) расходятся к полюсам клетки, В конце профазы начинает образовываться веретено деления, которое формируется из микротрубочек.
В метафазе завершается образование веретена деления, которое состоит из микротрубочек двух типов: хромосомных, которые с двух полюсов связываются с центромерами хромосом, и центросомных (полюсных), которые тянутся от полюса к полюсу клетки. Хромосомы как бы выталкиваются микротрубочками в область экватора клетки. В метафазе отчетливо видно двойное строение хромосом, соединенных только в области центромеры. В этот период легко подсчитывать число хромосом, изучать их особенности морфологического строения.
В анафазе дочерние хромосомы (хроматиды) с помощью микротрубочек веретена деления расходятся и движутся к полюсам клетки. Во время движения дочерние хромосомы несколько изгибаются наподобие шпильки, концы которой повернуты в сторону экватора клетки. Таким образом, в анафазе хроматиды удвоенных в интерфазе хромосом расходятся к полюсам клетки. В этот момент в клетке находятся два диплоидных набора хромосом.
В телофазе происходят процессы, обратные тем, которые наблюдаются в профазе: начинается деспирализация (раскручивание) хромосом, они набухают и становятся плохо различимы в световом микроскопе, Вокруг хромосом у каждого полюса из мембранных структур цитоплазмы формируется ядерная оболочка, в ядрах возникают ядрышки. Разрушается веретено деления.
На стадии телофазы происходит разделение цитоплазмы с образованием двух клеток.
В клетках животных цитоплазма делится путем кольцевидной перетяжки с образованием двух отдельных клеток.
В клетках растений в области экватора из остатков нитей веретена деления возникает бочковидное образование— фрагмо-пласт. В эту область со стороны полюсов клетки устремляются многочисленные пузырьки комплекса Гольджи, которые сливаются друг с другом. Содержимое пузырьков образует срединную пластинку, которая делит клетку на две дочерние, а мембрана пузырьков образует недостающие цитоплазматические мембраны этих клеток. Впоследствии на срединную пластинку со стороны каждой из дочерних клеток откладываются элементы клеточных оболочек.
В результате митоза из одной клетки возникают две дочерние с тем же набором хромосом.
Таким образом, биологический смысл митоза состоит в строго одинаковом распределении между дочерними клетками материальных носителей наследственности — молекул ДНК, входящих в состав хромосом материнской клетки. Благодаря равномерному разделению реплицированных хромосом между дочерними клетками обеспечивается образование генетически равноценных клеток и сохраняется преемственность в ряду клеточных поколений. Это обеспечивает такие важные моменты жизнедеятельности, как эмбриональное развитие и рост организмов, восстановление органов и тканей после повреждения. Митотическое деление клеток является цитологической основой бесполого размножения организмов.
Бактериальные клетки содержат только одну кольцевую молекулу ДНК, прикрепленную к клеточной мембране. Перед делением клетки ДНК реплицируется и образуются две идентичные молекулы ДНК, каждая из которых также прикреплена к клеточной мембране.
При делении клетки мембрана врастает между двумя молекулами ДНК так, что в конечном итоге в каждой дочерней клетке оказывается по одной идентичной молекуле ДНК. Такой процесс получил название прямого бинарного деления.
Амитоз или прямое деление, — это деление интерфазного ядра путем перетяжки. При амитозе веретено деления не образуется и хромосомы в световом микроскопе неразличимы. Такое деление встречается у одноклеточных организмов (например, так делятся большие полиплоидные ядра инфузорий), а также в некоторых высокоспециализированных с ослабленной физиологической активностью, дегенерирующих, обреченных на гибель клетках растений и животных либо при различных патологических процессах, таких как злокачественный рост, воспаление и т. п.
Амитоз можно наблюдать в тканях растущего клубня картофеля, эндосперме семян, стенках завязи пестика и паренхиме черешков листьев. У животных и человека такой тип деления характерен для клеток печени, хрящей, роговицы глаза.
При амитозе часто наблюдается только деление ядра: в этом случае могут возникнуть двух- и многоядерные клетки. Если же за делением ядра следует деление цитоплазмы, то распределение клеточных компонентов, как и ДНК, осуществляется произвольно.
Амитоз в отличие от митоза является самым экономичным способом деления, так как энергетические затраты при этом весьма незначительны.
Мейоз (греч. meiosis -— уменьшение) — это особый способ деления клеток, в результате которого происходит редукция (уменьшение) числа хромосом вдвое и переход клеток из диплоидного состояния (2л) в гаплоидное (п) (рис. 1.25).
С помощью мейоза образуются споры высших растений и половые клетки —гаметы, В результате редукции хромосомного набора в каждую гаплоидную спору и гамету попадает по одной хромосоме из каждой пары хромосом, имеющихся в данной клетке. В ходе дальнейшего процесса оплодотворения (слияния гамет) организм нового поколения получит также диплоидный набор хромосом, т. е. кариотип организмов данного вида в ряду поколений остается постоянным. Важнейшее значение мейоза заключается в обеспечении постоянства кариотипа в ряду поколений организмов данного вида.

Рис. 1.25. Схема мейоза (репликация хромосомного материала происходит в S-nepuoдe интерфазы),
Мейоз включает два быстро следующих одно за другим деления. Перед началом мейоза каждая хромосома реплицируется. В течение некоторого времени две ее образовавшиеся копии остаются связанными друг с другом центромерой. Следовательно, в каждом ядре, в котором начинается мейоз, содержится эквивалент четырех наборов гомологичных хромосом 4с (с — сестринские хроматиды — по две в каждой гомологичной хромосоме). Поэтому для образования ядра гамет, содержащих гаплоидный набор хромосом, необходимо два ядерных деления. Эти деления так и называются: первое деление мейоза и второе деление мейоза. Второе деление мейоза следует практически сразу же за первым, и синтеза ДНК в промежутке между ними не происходит (т. е. между первым и вторым делением отсутствует интерфаза).
Первое мейотическое (редукционное) деление приводит к образованию из диплоидных клеток (2n) гаплоидных клеток (п). Оно начинается с профазы I, в которой, как и в митозе, осуществляется упаковка наследственного материала (спиралазация хромосом). Одновременно происходит конъюгация — сближение гомологичных (парных) хромосом — соединяются одинаковые участки (в митозе не наблюдается). В результате конъюгации образуются хромосомные пары — биваленты (рис. 1.25). Каждая хромосома, вступая в мейоз, как уже отмечалось, имеет удвоенное количество наследственного материала и состоит из двух хро-матид, поэтому бивалент включает уже четыре нити.
Когда хромосомы находятся в конъюгированном состоянии, продолжается их дальнейшая спирализация. При этом отдельные хроматиды гомологичных хромосом переплетаются между собой. В последующем гомологичные хромосомы отталкиваются и несколько отходят одна от другой. В результате этого в местах переплетения хроматид может происходить их разрыв, и, как следствие, в процессе воссоединения этих разрывов гомологичные хромосомы обмениваются соответствующими участками. В результате хромосома, перешедшая к данному организму от отца, включает участок материнской хромосомы, и наоборот. Перекрест гомологичных хромосом, сопровождающийся обменом соответствующими участками между их хро-матидами, называется кроссинговером. После кроссинговера расходятся уже измененные хромосомы, т. е. с другим сочетанием генов. Являясь процессом закономерным, кроссинговер приводит каждый раз к обмену разными по величине участками и обеспечивает эффективную рекомбинацию материала хромосом в гаметах.
К концу профазы хромосомы в бивалентах, сильно спирали-зуясь, укорачиваются. Как и в митозе, в конце профазы I разрушается ядерная оболочка и начинает формироваться веретено деления.
В метафазе I завершается формирование веретена деления. Его нити прикрепляются к центромерам хромосом, объединенных в биваленты таким образом, что к каждой центромере идет лишь одна нить от одного из полюсов клетки. В результате нити, связанные с центромерами гомологичных хромосом, устанавливают биваленты в плоскости экватора веретена деления.
В анафазе I гомологичные хромосомы, каждая из которых состоит из двух хроматид, разделяются и расходятся к полюсам клетки.
В телофазе I у полюсов веретена деления собирается половинное число хромосом (гаплоидный набор). В этой короткой по продолжительности фазе восстанавливается ядерная оболочка, после чего материнская клетка делится на две дочерние.
Таким образом, образование бивалентов при конъюгации гомологичных хромосом в профазе I мейоза создает условия для последующей редукции числа хромосом. Формирование их гаплоидного набора в гаметах обеспечивается расхождением в анафазе 1 не хроматид, как в митозе, а гомологичных хромосом, которые ранее были объединены в биваленты.
Второе мейотическое деление следует сразу за первым и сходно с обычным митозом (поэтому его часто называют митозом мейоза), но в отличие от митоза клетки, вступающие в него, имеют гаплоидный набор хромосом.
Профаза II непродолжительна. В метафазе II снова образуется веретено деления, хромосомы выстраиваются в экваториальной плоскости и центромерами прикрепляются к микротрубочкам веретена деления. В анафазе II осуществляется разделение их центромер, и каждая хроматида становится самостоятельной хромосомой. Отделившиеся друг от друга дочерние хромосомы растягиваются микротрубочками веретена деления к полюсам. В телофазе II завершается расхождение сестринских хромосом к полюсам и наступает деление клеток: из двух гаплоидных клеток образуются четыре гаплоидные дочерние клетки. Таким образом, в результате мейоза из одной диплоидной клетки образуются четыре клетки с гаплоидным набором хромосом.
Редукционное деление является как бы регулятором, препятствующим непрерывному увеличению числа хромосом при слиянии гамет. Не будь такого механизма, при половом размножении число хромосом удваивалось бы в каждом новом поколении.
Иными словами, благодаря мейозу поддерживается определен-ное и постоянное число хромосом во всех поколениях каждого вида растений, животных и грибов.
Другое важное значение мейоза заключается в обеспечении чрезвычайного разнообразия генетического состава гамет в результате как кроссинговера, так и различного сочетания отцовских и материнских хромосом при их расхождении в анафазе I. Это обеспечивает появление разнообразного и разнокачественного потомства при половом размножении организмов.