Пластический обмен
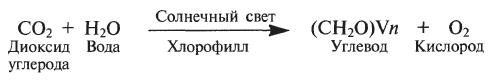
Фотосинтез. Это процесс преобразования энергии света в энергию химических связей органических веществ. Процесс фотосинтеза обычно описывают уравнением:
Такое преобразование происходит в хлоропластах, где имеются молекулы хлорофиллов, поглощающих световые волны разной длины. Важнейшими из них являются хлорофиллы П700 и П680, поглощающие свет с длиной волны 700 и 680 нм соответственно. Они входит в состав реакционных центров фотосистем I и II.
Процесс фотосинтеза представляет собой цель окислительно-восстановительных реакций, где происходит восстановление углекислого газа до органических веществ. Всю совокупность фотосинтетических реакций принято подразделять на две фазы— световую и темновую. Темновая фаза происходит параллельно световой с использованием продуктов, образованных в световой фазе.
Световая фаза фотосинтеза. Прохождение световой фазы связано с мембранами тилакоидов при участии хлорофилла и других пигментов, фермента АТФ-синтетазы, встроенного в мембрану тилакоидов, и белков-переносчиков.
Для световой фазы фотосинтеза характерно то, что энергия солнечной радиации, поглощенная хлорофиллами, преобразуется сначала в электрохимическую, а затем в энергию макроэрги-ческих связей АТФ. Это достигается путем переноса электронов и ионов водорода с помощью специальных переносчиков через мембрану тилакоидов (рис. 1.18).
Световая фаза фотосинтеза разделяется на фотофизическую и фотохимическую. В фотофизической фазе происходит поглощение квантов света молекулами хлорофиллов П700(фотосистема I) и П680 (фотосистема II) и переход этих молекул в возбужденное состояние.
В фотохимической фазе обе фотосистемы работают согласованно.
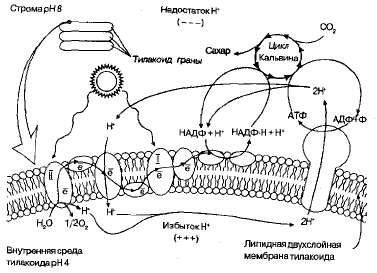
Рис 1.18. Схема фотосинтеза (стрелками показан перенос протонов и электронов).
Фотосистема I. Возбужденная молекула П700 отдает электрон акцептору. От него по системе переносчиков этот электрон попадает на внешнюю сторону тилакоида (обращенную в стро-му). При этом молекула П700 окисляется и П700 превращется в П700 +:
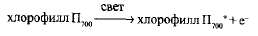
Фотосистема II. Возбужденная молекула П680 отдает электрон акцептору. От него по системе переносчиков электрон переходит в фотосистему I и восстанавливает молекулу П700. При этом молекула П70О возвращается в исходное состояние и становится вновь способной поглощать свет и переходить в возбужденное состояние.
Молекула же П680, отдав электрон, превращается в П680+. Для ее восстановления используются электроны, отщепляемые от молекул воды в процессе так называемого фотолиза,или фотоокисления:
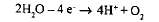
Благодаря фотолизу внутри тилакоида накапливаются протоны и образуется молекулярный кислород, который диффундирует в атмосферу. Кислород является побочным продуктом реакций фотосинтеза.
Протоны, образовавшиеся при фотолизе, не могут проникнуть через мембрану тилакоида и накапливаются внутри, создавая Н+-резервуар. Таким образом, по разные стороны мембраны накапливаются протоны и электроны, т. е. возникает электрохимический мембранный потенциал. Когда он достигает величины 200 мВ, протоны проталкиваются силой электрического поля через каналы фермента АТФ-синтетазы, встроенного в мембрану тилакоидов, т. е. начинает работать протонная помпа. На выходе из протонного канала создается высокий уровень энергии, которая используется для фосфорилирования имеющихся в матриксе хлоропластов молекул АДФ:
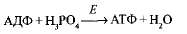
Перенесенные протоны взаимодействуют с электронами и молекулой НАДФ (кофермент никотинамидадениндинуклеотидфос-фат), образуя комплекс НАДФН + Н+:
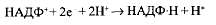
Таким образом, активированные световой энергией электроны используются для присоединения атома водорода к его переносчику, т. е. на восстановление НАДФ' в НАДФН + Н+
В конечном итоге в световой фазе фотосинтеза образуются АТФ, НАДФ-восстановленный и кислород, являющийся побочным продуктом фотосинтеза. Полученные АТФ и НАДФ-восстановленный используются в темновой фазе фотосинтеза.
Темновая фаза фотосинтеза. Этот сложный процесс, осуществляемый в строме хлоропластов без непосредственного поглощения света, включает большое количество реакций, приводящих к восстановлению С02 до уровня органических веществ, за счет использования энергии АТФ и НАДФ-Н + Н, синтезированных в световую фазу. Существуют разные пути восстановления. Основным из них является так называемый Сз-путь, или цикл Кальвина. Для синтеза в нем одной молекулы глюкозы необходимо 12 молекул НАДФ-Н + Н+ и 18 молекул АТФ, которые поставляются в результате фотохимических реакций фотосинтеза. СO2 поступа-ет в хлоропласты из атмосферы.
В темновой фазе фотосинтеза, таким образом, энергия макроэргических связей АТФ преобразуется в химическую энергию
органических веществ, т. е. энергия как бы консервируется в химических связях органических веществ.
Если реакции световой и темновой фаз объединить, исключив все промежуточные стадии и вещества, то можно получить суммарное уравнение процесса фотосинтеза, которое было приведено в начале параграфа.
Таким образом, фотосинтез —процесс, при котором происходит поглощение электромагнитной энергии Солнца хлорофиллом и вспомогательными пигментами, поглощение углекислого газа из атмосферы, восстановление его в органические соединения и выделение кислорода в атмосферу.
На скорость фотосинтеза оказывают влияние различные факторы окружающей среды: интенсивность падающего света, наличие влаги, минеральных веществ, температура, концентрация С02 и др.
Значение фотосинтеза. Уникальность и общебиологическое значение фотосинтеза определяются тем, что ему обязано своим существованием все живое на нашей планете. Этот процесс является основным источником образования первичных органических веществ, а также единственным источником свободного кислорода на Земле. Из кислорода образовался и поддерживается озоновый слой, защищающий живые организмы от воздействия коротковолновой ультрафиолетовой радиации. Кроме того, благодаря фотосинтезу регулируется содержание С02 в атмосфере.
В изучение процесса фотосинтеза, раскрытие его механизма большой вклад внесли русский ученый К. А. Тимирязев, американский М. Кальвин, австралийские М. Д. Хетч и К. Р. Слэк, а также наши белорусские ученые академики Т. Н. Годнев и А. А. Шлык.
Хемосинтез. Кроме фотосинтеза существует еще одна форма автотрофной ассимиляции — хемосинтез, свойственный некоторым бактериям. В отличие от фотосинтеза при хемосинтезе используется не световая энергия, а энергия, выделенная при окислении некоторых неорганических соединений, например сероводорода, серы, аммиака, водорода, азотистой кислоты, оксидных соединений железа и марганца и др.
Открытие бактериального хемосинтеза принадлежит русскому ученому-микробиологу С. Н. Виноградскому.
Важнейшей группой хемосинтезирующих организмов являются нитрифицирующие бактерии, способные окислять образующийся при гниении органических остатков аммиак до нитрита, а затем и до нитрата:
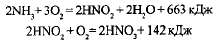
Азотная кислота, реагируя с минеральными соединениями почвы, превращается в соли азотной кислоты, которые хорошо усваиваются растениями.
Бесцветные серобактерии окисляют сероводород и накапливают в своих клетках серу
: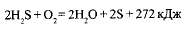
При недостатке сероводорода бактерии производят дальнейшее окисление накопившейся в них серы до серной кислоты
.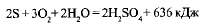
Железобактерии переводят железо Fe2+ в железо Fe3+
: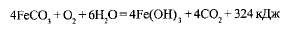
Водородные бактерии используют в качестве источника энергии реакции окисления молекулярного водорода, а в качестве единственного источника углерода — углекислый газ. Реакция окисления происходит по схеме:
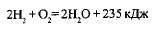
Энергия, выделяемая при окислении указанных выше соединений, используется бактериями-хемосинтетиками для восстановления С02 до органических веществ.
Хемотрофные нитрифицирующие бактерии широко распространены в природе. Они встречаются и в почве, и в разных водоемах. Масштаб осуществляемых ими процессов достаточно велик, поэтому хемосинтетики играют существенную роль в круговороте азота в биосфере.
Серобактерии способствуют постепенному разрушению и выветриванию горных пород вследствие образования ими серной кислоты, являются причиной порчи каменных и металлических сооружений, выщелачивания руд и серных месторождений. Многие виды серобактерий, окисляя до сульфатов различные соединения серы, играют большую роль в процессах очищения промышленных сточных вод.
В результате деятельности некоторых железобактерий вырабатывается Fe(OH)3, скопления которого образуют болотную железную руду. Водородные бактерии используются для получения дешевого пищевого и кормового белка. Кроме того, в природных условиях водородные бактерии участвуют в окислении водорода, который может накапливаться при действии некоторых микроорганизмов, размельчающих органические вещества почвы, донные отложения водоемов и т. д.
Биосинтез белка. Важнейшим анаболическим процессом является синтез белка. Все морфологические и функциональные особенности любой клетки и организма в целом определяются структурой специфических белков, входящих в состав клеток. Способность к синтезу только строго определенных белков является наследственным свойством организмов. Последовательность расположения аминокислот в полипептидной цепи (первичной структуре белка), от которой зависят его биологические свойства, определяется последовательностью нуклеотидов в молекуле ДНК.
Последовательность нуклеотидов в полинуклеотидной цепи специфична для каждой клетки и представляет собой генетический код, посредством которого записана информация о синтезе белков. Это значит, что в ДНК каждое сообщение закодировано специфической последовательностью из четырех знаков — А, Г, Т, Ц, подобно тому как письменное сообщение кодируется знаками (буквами) алфавита или азбуки Морзе.
Генетический код характеризуется следующими свойствами:
- Код является триплетным, т.е. каждая аминокислота кодируется известным сочетанием из трех последовательно расположенных нуклеотидов, которое называется триплетомили кодонам. Нетрудно подсчитать, что число возможных комбинаций из четырех нуклеотидов по три составит 64, что более чем достаточно для кодирования 20 аминокислот, входящих в состав белка.
- Код является множественным, или «вырожденным», т. е. одна и та же аминокислота может кодироваться несколькими триплетами (от 2 до 6), в то время как каждый триплет кодирует только одну аминокислоту, например:
фенилаланин — AAA, ААГ;
изолейцин — ТАА, ТАГ, ТАТ;
пролин — ГГА, ГГГ, ГГТ, ГГЦ;
серин — АГА, АГГ, АГТ, АГЦ, ТЦА, ТЦГ.
Исключение составляют метионин и триптофан, которые кодируются только одним триплетом (ТАЦ и АЦЦ в ДНК и соответственно АУГ и УГГ в РНК). - Код является неперекрывающимся, т. е. один и тот же нуклео-тид не может входить одновременно в состав двух соседних триплетов.
- Код однозначен, т. е. каждый триплет кодирует только одну аминокислоту.
- Код не имеет знаков препинания. Это значит, что если произойдет выпадение одного нуклеотида, то при считывании его место займет ближайший нуклеотид из соседнего кодона, из-за чего изменится весь порядок считывания. Вместе с тем между генами имеются триплеты, обозначающие прекращение синтеза одной гюлипептидной цепи (в ДНК это АТТ, АТЦ и АЦТ, а в РНК соответственно УАА, УАГ и УГА).
- Код универсален для всех живых организмов и вирусов: одинаковые триплеты кодируют одинаковые аминокислоты. Это открытие представляет собой серьезный шаг на пути к более глубокому познанию сущности живой материи, ибо универсальность генетического кода свидетельствует о единстве происхождения всех живых организмов.
К настоящему времени расшифрованы триплеты для всех 20 аминокислот, входящих в состав природных белков. Зная порядок расположения триплетов в молекуле ДНК (генетический код), можно установить последовательность расположения аминокислот в белке. В одной молекуле ДНК может быть закодирована последовательность аминокислот для многих белков. Функциональный отрезок молекулы ДНК, несущий в себе информацию о структуре одного белка, называется геном. Различают структурные гены, в которых закодирована информация для синтеза структурных и ферментных белков, и гены с информацией для синтеза тРНК, рРНК.
Итак, план построения белка закодирован в ДНК, которая непосредственного участия в синтезе белковых молекул не принимает.
Процесс биосинтеза белка осуществляется на рибосомах, расположенных преимущественно в цитоплазме. Следовательно, для передачи генетической информации с ядерной ДНК к месту синтеза белка требуется посредник. Таким посредником является иРНК, которая на основе принципа комплементарности синтезируется на одной из цепей молекулы ДНК. Этот процесс получил название транскрипции или переписывания.
Транскрипция происходит не на всей молекуле ДНК одновременно, а лишь на небольшом ее участке, отвечающем определенному гену (рис. 1.19). При этом часть двойной спирали ДНК раскручивается, обнажая короткий участок одной из цепей, который теперь будет служить матрицей для синтеза иРНК. Вдоль этой цепи движется фермент РНК-полимераза, соединяя между собой нуклеотиды в растущую цепь иРНК, последовательность нуклеотидов которой является точной копией последовательности нуклеотидов матрицы — одного или группы рядом расположенных генов. Так, если в молекуле ДНК имеется азотистое основание цитозин, то в РНК — гуанин, и наоборот. В ДНК комплементарной парой являются аденин и тимин. Однако в состав иРНК вместо тимина входит урацил. Транскрипция может происходить одновременно на нескольких генах одной хромосомы и на генах, расположенных на разных хромосомах.
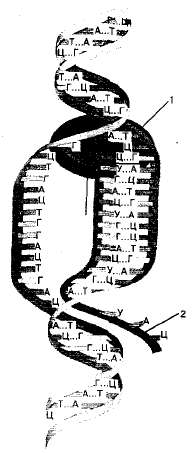
Рис. 1.19. Схема транскрипции: 1 — РНК-полимераза; 2 — синтезируемая молекула иРНК.
На специальных генах синтезируются и два других типа РНК — тРНК и рРНК. Начало и конец синтеза всех типов РНК на матрице ДНК строго фиксирован специальными триплетами, которые контролируют запуск (инициирующие) и остановку (терминирующие) синтеза РНК. Триплеты выполняют функцию знаков препинания между генами.
Синтезированная в ядре иРНК отделяется от ДНК и через поры ядерной оболочки поступает в цитоплазму, где прикрепляется к рибосоме.
Молекула иРНК может связываться одновременно с несколькими рибосомами. Их число определяется длиной иРНК. Комплекс из иРНК и рибосом (от 5-6 до нескольких десятков) называется полирибосомой или полисомой. Образование полисом повышает эффективность функционирования иРНК за счет того, что одновременно протекает синтез нескольких идентичных полипептидных цепей. Именно на полисомах происходит синтез белка, или трансляция (рис. 1.20).
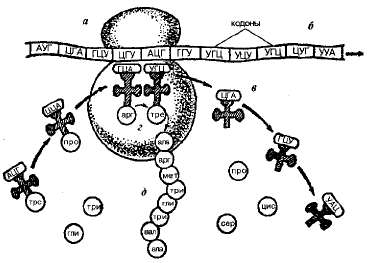
Рис 1.20. Схема биосинтеза белка: а — рибосома; б — иРНК; в — тРНК; г — образование пептидной связи; д —растущая полипептид ноя цепь.
Трансляция начинается со стартового кодона АУГ. Отсюда молекула иРНК прерывисто, триплет за триплетом продвигается через рибосомы, что сопровождается ростом полипептидной цепочки. Число аминокислот в таком белке равно числу триплетов иРНК.
Выстраивание аминокислот в соответствии с кодонами иРНК осуществляется на рибосомах при помощи тРНК — главных агентов синтеза белка. Благодаря определенному расположению комплементарных нуклеотидов цепочка тРНК имеет форму, напоминающую лист клевера (рис. 1.21). При этом тРНК имеет акцепторный конец, к которому присоединяется активированная энергией АТФ аминокислота.
Активацию аминокислот осуществляют специфичные ферменты аминоацил-тРНК-синтетазы, т. е. для каждой аминокислоты существует свой фермент. Механизм активации заключается в том, что фермент одновременно взаимодействует с соответствующей аминокислотой и АТФ, которая теряет при этом пирофосфат. Тройной комплекс из фермента, аминокислоты и АТФ называется активированной (богатой энергией) аминокислотой, которая способна спонтанно образовать пептидную связь, что приводит к синтезу полипептидов. Данный процесс активации — необходимый этап белкового синтеза, поскольку свободные аминокислоты не могут прямо присоединяться к полипептидной цепи.
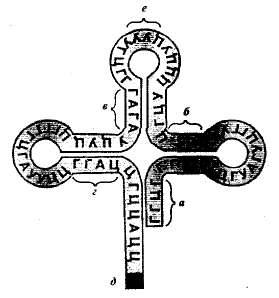
Рис 1.21. Структура тРНК: а, б, в, г — участки комплементарного соединения; д —участок соединения с аминокислотой; е—антикодон.
В противоположной части молекулы тРНК располагается специфический триплет (антикодон), ответственный за прикрепление по принципу комплементарности к определенному триплету иРНК (кодой).
тРНК с присоединенной к ней активированной аминокислотой антикодоном присоединяется к кодону иРНК. Затем к той же рибосоме прикрепляется вторая тРНК с аминокислотой в соответствии со следующим кодоном. В функциональном центре рибосомы оказываются два триплета (кодона), с которыми взаимодействуют две транспортные РНК. В рибосоме, таким образом, оказались две аминокислоты, между которыми возникает пептидная связь. Первая тРНК, освободившись от аминокислоты, покидает рибосому. Далее к образованному дипептиду аналогичным путем пристраивается третья, четвертая и т. д. аминокислоты, принесенные в рибосому своими тРНК. Процесс продолжается до тех пор, пока в рибосому не попадет один из трех терминирующих кодонов: УАА, УАГ или УГА, — после чего синтез белка прекращается.
Таким образом, последовательность кодонов иРНК определяет последовательность включения аминокислот в цепь белка. После завершения синтеза белка иРНК под действием ферментов распадается на отдельные нуклеотиды.
Таким образом, роль нуклеиновых кислот в биосинтезе белка заключается в преобразовании генетической информации, представленной в виде последовательности нуклеотидов ДНК, в структуру молекулы иРНК, а затем в последовательность аминокислот в молекуле белка. Реакции синтеза иРНК (транскрипция) и белка (трансляция) осуществляются по матрицам (ДНК и иРНК соответственно), поэтому они получили название реакций матричного синтеза.
Каждый этап биосинтеза белка катализируется соответствующими ферментами и снабжается энергией за счет расщепления АТФ. Синтезированные белки поступают в каналы эндоплазматической сети, где происходит их дозревание, приобретение вторичной, третичной и четвертичной структур.
Синтез белковых молекул происходит непрерывно и идет с большой скоростью: за 1 мин образуется от 50 до 60 тыс. пептидных связей. Синтез одной молекулы белка длится всего 3—4 с. В результате половина белков тела человека (в среднем в нем около 17 кг белка) обновляется за 80 суток. За всю жизнь в организме человека белки обновляются около 200 раз.