Органеллы клетки и их функции
Основные группы органелл. Органеллы — постоянные внутриклеточные структуры, имеющие определенное строение и выполняющие соответствующие функции. Органеллы делятся на две группы: мембранные и немембранные. Мембранные органеллы представлены двумя вариантами: двумембранным и одномем-бранным. Двумембранными компонентами являются пластиды, митохондрии и клеточное ядро. К одномембранным относятся органеллы вакуолярной системы — эндоплазматический рети-кулум, комплекс Гольджи, лизосомы, вакуоли растительных и грибных клеток, пульсирующие вакуоли и др. К немембранным органеллам принадлежат рибосомы и клеточный центр, постоянно присутствующие в клетке. Выраженность элементов цитоскелета (постоянного компонента клетки) может значительно меняться в течение клеточного цикла — от полного исчезновения одного компонента (например, цитоплазматических трубочек во время деления клетки) до появления новых структур (веретена деления).
Общим свойством мембранных органелл является то, что все они построены из липопротеидных пленок (биологических мембран), замыкающихся сами на себя так, что образуются замкнутые полости, или отсеки. Внутреннее содержимое этих отсеков всегда отличается от гиалоплазмы.
Двумембранные органеллы. К двумебранным органеллам относятся пластиды и митохондрии. Пластиды —характерные органеллы клеток автотрофных эукариотических организмов. Их окраска, форма и размеры весьма разнообразны. Различают хлоропласты, хромопласты и лейкопласты.
Хлоропласты имеют зеленый цвет, обусловленный присутствием основного пигмента — хлорофилла. Хлоропласты содержат также вспомогательные пигменты — каротиноиды (оранжевого цвета). По форме хлоропласты — это овальные линзовидные тельца размером (5—10) х (2—4) мкм. В одной клетке листа может находиться 15—20 и более хлоропластов, а у некоторых водорослей — лишь 1 -2 гигантских хлоропласта (хроматофора) различной формы.
Хлоропласты ограничены двумя мембранами — наружной и внутренней (рис. 1.8).
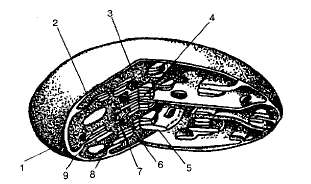
Рис. 1.8. Схема строения хлоропласта: I —наружная мембрана; 2 — рибосомы; 3 — пластоглобулы; 4 — граны; 5 — тилакоиды; 6 — матрице; 7 —ДНК; 8 — внутренняя мембрана; 9 —межмембранное пространство.
Наружная мембрана отграничивает жидкую внутреннюю гомогенную среду хлоропласта — строму (матрикс). В строме содержатся белки, липиды, ДНК (кольцевая молекула), РНК, рибосомы и запасные вещества (липиды, крахмальные и белковые зерна) а также ферменты, участвующие в фиксации углекислого газа.
Внутренняя мембрана хлоропласта образует впячивания внутрь стромы —тилакоиды, или ламеллы, которые имеют форму уплощенных мешочков (цистерн). Несколько таких тилакоидов, лежащих друг над другом, образуют грану, и в этом случае они называются тилакоидами граны. Именно в мембранах тилакоидов локализованы светочувствительные пигменты, а также переносчики электронов и протонов, которые участвуют в поглощении и преобразовании энергии света.
Хлоропласты в клетке осуществляют процесс фотосинтеза.
Лейкопласты — мелкие бесцветные пластиды различной формы. Они бывают шаровидными, эллипсоидными, гантелевидными, чашевидными и т. д. По сравнению с хлоропластами у них слабо развита внутренняя мембранная система.
Лейкопласты в основном встречаются в клетках органов, скрытых от солнечного света (корней, корневищ, клубней, семян). Они осуществляют вторичный синтез и накопление запасных питательных веществ — крахмала, реже жиров и белков.
Хромопласты отличаются от других пластид своеобразной формой (дисковидной, зубчатой, серповидной, треугольной, ромбической и др.) и окраской (оранжевые, желтые, красные). Хромопласты лишены хлорофилла и поэтому не способны к фотосинтезу. Внутренняя мембранная структура их слабо выражена.
Хромопласты присутствуют в клетках лепестков многих растений (лютиков, калужниц, нарциссов, одуванчиков и др.), зрелых плодов (томаты, рябина, ландыш, шиповник) и корнеплодов (морковь, свекла), а также листьев в осеннюю пору. Яркий цвет этих органов обусловлен различными пигментами, относящимися к группе каргиноидов, которые сосредоточены в хромопластах.
Все типы пластид генетически родственны друг другу, и одни их виды могут превращаться в другие:
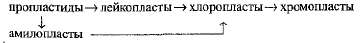
Таким образом, весь процесс взаимопревращений пластид можно представить в виде ряда изменений, идущих в одном направлении — от пропластид до хромопластов.
Митохондрии—неотъемлемые компоненты всех эукариотических клеток. Они представляют собой гранулярные или нитеподобные структуры толщиной 0,5 мкм и длиной до 7—10 мкм.
Митохондрии ограничены двумя мембранами — наружной и внутренней (рис. 1.9). Между внешней и внутренней мембранами имеется так называемое перимитохондриалъное пространство, которое является местом скопления ионов водорода Н+ Наружная митохондриальная мембрана отделяет ее от гиалоплазмы. Внутренняя мембрана образует множество впячиваний внутрь митохондрий — так называемых крист. На мембране крист или внутри нее располагаются ферменты, в том числе переносчики электронов и ионов водорода Н+, которые участвуют в кислородном дыхании. Наружная мембрана отличается высокой проницаемостью, и многие соединения легко проходят через нее. Внутренняя мембрана менее проницаема. Ограниченное ею внутреннее содержимое митохондрии {матрикс) по составу близко к цитоплазме. Матрикс содержит различные белки, в том числе ферменты, ДНК (кольцевая молекула), все типы РНК, аминокислоты, рибосомы, ряд витаминов. ДНК обеспечивает некоторую генетическую автономность митохондрий, хотя в целом их работа координируется ДНК ядра.
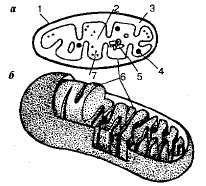
Рис. 1.9. Схема строения митохондрии: а — продольный разрез; 6 — схема трехмерного строения; 1 — внешняя мембрана; 2 — матрикс; 3 —межмембранное пространство; 4— гранула; 5 —ДНК; 6 — внутренняя мембрана; 7 — рибосомы.
В митохондриях осуществляется кислородный этап клеточного дыхания.
Одномембранные органеллы. В клетке синтезируется огромное количество различных веществ. Часть из них потребляется на собственные нужды (синтез АТФ, построение органелл, накопление питательных веществ), часть выводится из клетки и используется на построение оболочки (клетки растений и грибов), гликокаликса (животные клетки). Клеточными секретами являются также ферменты, гормоны, коллаген, кератин и т. д. Накопление этих веществ и перемещение их из одной части клетки в другую либо выведение за ее пределы происходит в системе замкнутых цитоплазматических мембран — эндоплазматической сети, или эндоплазматическом ретикулуме, и комплексе Гольджи, составляющих транспортную систему клеток.
Эндоплазматический ретикулум был открыт с помощью электронного микроскопа в 1945 г. Он представляет собой систему разветвленных каналов, цистерн (вакуолей), пузырьков, создающих подобие рыхлой сети в цитоплазме (рис. 1.10). Стенки каналов и полостей образованы элементарными мембранами.
В клетке существует два типа эндоплазматического ретикулума: гранулярный (шероховатый) и агранулярный (гладкий). Гранулярный эндоплазматический ретикулум густо усеян рибосомами, на которых осуществляется биосинтез белка. Синтезируемые белки проходят через мембрану в каналы и полости эндоплазматического ретикулума, изолируются от цитоплазмы, накапливаются там, дозревают и перемещаются в другие части клетки либо в комплекс Гольджи в специальных мембранных пузырьках, которые отшнуровываются от цистерн эндоплазматического ретикулума.
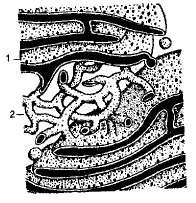
Рис. 1.10. Схема строения шероховатого (1) и гладкого (2) эндоплазматического ретикулума.
Функции эндоплазматического ретикулума следующие:
- В мембранах гранулярного эндоплазматического ретикулума накапливаются и изолируются белки, которые после их синтеза могли оказаться вредными для клетки. Например, синтез гидролитических ферментов и их свободный выход в цитоплазму привел бы к самоперевариванию клетки и ее гибели. Однако этого не происходит, потому что подобные белки надежно изолированы в полостях эндоплазматического ретикулума.
- На рибосомах гранулярного эндоплазматического ретикулума синтезируются также интегральные и периферические белки мембран клетки и некоторая часть белков цитоплазмы.
- Цистерны шероховатого эндоплазматического ретикулума связаны с ядерной оболочкой, причем некоторые из них являются прямым продолжением последней. Считается, что после деления клетки оболочки новых ядер образуются из цистерн эндоплазматического ретикулума.
- На мембранах гладкого эндоплазматического ретикулума протекают процессы синтеза липидов и некоторых углеводов (например, гликогена).
Комплекс (аппарат) Голъджи открыт в 1898 г. итальянским ученым К. Гольджи. Он представляет собой систему плоских дисковидных замкнутых цистерн, которые располагаются одна над другой в виде стопки и образуют диктиосому. От цистерн отходят во все стороны мембранные трубочки и пузырьки (рис. 1.11). Число диктиосом в клетках варьирует от одной до нескольких десятков в зависимости от типа клеток и фазы их развития.

Рис 1.11. Схема строения аппарата Голъджи: 1 — пузырьки; 2 — цистерны.
К комплексу Гольджи доставляются вещества, синтезируемые в эндоплазматическом ретикулуме. От цистерн эндоплазматического ретикулума отшнуровываются пузырьки, которые соединяются с цистернами комплекса Гольджи, где эти вещества модифицируются и дозревают.
Пузырьки комплекса Гольджи участвуют в формировании цитоплазматической мембраны и стенок клеток растений после деления, а также в образовании вакуолей и первичных лизосом.
Зрелые цистерны диктиосомы отшнуровывают пузырьки или вакуоли Гольджи, заполненные секретом. Содержимое таких пузырьков либо используется самой клеткой, либо выводится за ее пределы. В последнем случае пузырьки Гольджи подходят к плазматической мембране, соединяются с ней и изливают свое содержимое наружу, а их мембрана включается в плазматическую мембрану и таким образом происходит ее обновление.
Цистерны комплекса Гольджи активно извлекают моносахариды из цитоплазмы и синтезируют из них более сложные олиго- и полисахариды. У растений в результате этого образуются пектиновые вещества, гемицеллюлоза и целлюлоза, используемые для построения клеточной стенки, слизь корневого чехлика. У животных подобным образом синтезируются гликопротеины и гликолипиды гликокаликса, вырабатываются секрет поджелудочной железы, амилаза слюны, пептидные гормоны гипофиза, коллаген.
Комплекс Гольджи участвует в образовании лизосом, белков молока в молочных железах, желчи в печени, веществ хрусталика, зубной эмали и г. п.
Комплекс Гольджи и эндоплазматический ретикулум тесно связаны между собой; их совместная деятельность обеспечивает синтез и преобразование веществ в клетке, их изоляцию, накопление и транспорт.
Лизосомы — это мембранные пузырьки величиной до 2 мкм. Внутри лизосом содержатся гидролитические ферменты, способные переваривать белки, липиды, углеводы, нуклеиновые кислоты. Лизосомы образуются из пузырьков, отделяющихся от комплекса Гольджи, причем предварительно на шероховатом эндоплазматическом ретикулуме синтезируются гидролитические ферменты.
Сливаясь с эндоцитозными пузырьками, лизосомы образуют пищеварительную вакуоль (вторичная лизосома), где происходит расщепление органических веществ до составляющих их мономеров. Последние через мембрану пищеварительной вакуоли поступают в цитоплазму клетки. Именно так происходит, например, обезвреживание бактерий в клетках крови — нейтрофилах.
Вторичные лизосомы, в которых закончился процесс переваривания, практически не содержат ферментов. В них находятся лишь непереваренные остатки, т. е. негидролизуемый материал, который либо выводится за пределы клетки, либо накапливается в цитоплазме.
Расщепление лизосомами чужеродного, поступившего путем эндоцитоза материала называется гетерофагией. Лизосомы участвуют также в разрушении материалов клетки, например запасных питательных веществ, а также макромолекул и целых органелл, утративших функциональную активность (аутофагия). При патологических изменениях в клетке или ее старении мембраны лизосом могут разрушаться: ферменты выходят в цитоплазму, и осуществляется самопереваривание клетки —автолиз. Иногда с помощью лизосом уничтожаются целые комплексы клеток и органы. Например, когда головастик превращается в лягушку, лизосомы, находящиеся в клетках хвоста, переваривают его: хвост исчезает, а образовавшиеся во время этого процесса вещества всасываются и используются другими клетками тела.
Вакуоли — крупные мембранные пузырьки или полости в цитоплазме, заполненные клеточным соком. Вакуоли образуются в клетках растений и грибов из пузыревидных расширений эндоплазматического ретикулума или из пузырьков комплекса Гольджи. В меристематических клетках растений вначале возникает много мелких вакуолей. Увеличиваясь, они сливаются в центральную вакуоль, которая занимает до 70—90% объема клетки и может быть пронизана тяжами цитоплазмы (рис. 1.12).
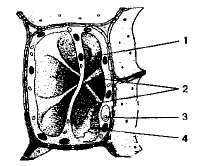
Рис. 1.12. Вакуоль в растительной клетке: 1 — вакуоль; 2 — цитопяаз-матические тяжи; 3 — ядро; 4 — хлоропласты.
Содержимое вакуолей —клеточный сок. Он представляет собой водный раствор различных неорганических и органических веществ. Большинство из них являются продуктами метаболизма протопласта, которые могут появляться и исчезать в различные периоды жизни клетки. Химический состав и концентрация клеточного сока очень изменчивы и зависят от вида растений, органа, ткани и состояния клетки. В клеточном соке содержатся соли, сахара (прежде всего сахароза, глюкоза, фруктоза), органические кислоты (яблочная, лимонная, щавелевая, уксусная и др.), аминокислоты, белки. Эти вещества являются промежуточными продуктами метаболизма, временно выведенными из обмена веществ клетки в вакуоль. Они являются запасными веществами клетки.
Помимо запасных веществ, которые могут вторично использоваться в метаболизме, клеточный сок содержит фенолы, танины (дубильные вещества), алкалоиды, антоцианы, которые выводятся из обмена в вакуоль и таким путем изолируются от цитоплазмы.
Танины особенно часто встречаются в клеточном соке (а также в цитоплазме и оболочках) клеток листьев, коры, древесины, незрелых плодов и семенных оболочек. Алкалоиды присутствуют, например, в семенах кофе (кофеин), плодах мака (морфин) и белены (атропин), стеблях и листьях люпина (люпинин) и др. Считается, что танины с их вяжущим вкусом, алкалоиды и токсичные полифенолы выполняют защитную функцию: их ядовитый (чаще горький) вкус и неприятный запах отталкивают растительноядных животных, что предотвращает поедание этих растений.
В вакуолях также часто накапливаются конечные продукты жизнедеятельности клеток (отходы). Таким веществом для клеток растений является щавелевокислый кальций, который откладывается в вакуолях в виде кристаллов различной формы.
В клеточном соке многих растений содержатся пигменты, придающие клеточному соку разнообразную окраску. Пигменты и определяют окраску венчиков цветков, плодов, почек и листьев, а также корнеплодов некоторых растений (например, свеклы).
Клеточный сок некоторых растений содержит физиологически активные вещества — фитогормоны (регуляторы роста), фитонциды, ферменты. В последнем случае вакуоли действуют как лизосомы. После гибели клетки мембрана вакуоли теряет избирательную проницаемость, и ферменты, высвобождаясь из нее, вызывают автолиз клетки.
Функции вакуолей следующие:
- Вакуоли играют главную роль в поглощении воды растительными клетками. Вода путем осмоса через ее мембрану поступает в вакуоль, клеточный сок которой является более концентрированным, чем цитоплазма, и оказывает давление на цитоплазму, а следовательно, и на оболочку клетки. В результате в клетке развивается тургорное давление, определяющее относительную жесткость растительных клеток и обусловливающее растяжение клеток во время их роста.
- В запасающих тканях растений вместо одной центральной часто бывает несколько вакуолей, в которых скапливаются запасные питательные вещества (жиры, белки).Сократительные (пульсирующие) вакуоли служат для осмотической регуляции, прежде всего, у пресноводных простейших, так как в их клетки путем осмоса непрерывно поступает вода из окружающего гипотонического раствора (концентрация веществ в речной или озерной воде значительно ниже, чем концентрация веществ в клетках простейших). Сократительные вакуоли поглощают избыток воды и затем выводят ее наружу путем сокращений.
Немембранные органеллы. Клеточный центр. В клетках большинства животных, а также некоторых грибов, водорослей, мхов и папоротников имеются центриоли. Расположены они обычно в центре клетки, что и определило их название (рис .1.13).

Центриоли представляют собой полые цилиндры длиной не более 0,5 мкм. Они располагаются парами перпендикулярно одна к другой (рис. 1.14). Каждая центриоль построена из девяти триплетов микротрубочек.
Основная функция центриолей — организация микротрубочек веретена деления клетки.
Центриолям по структуре идентичны базальные тельца, которые всегда обнаруживаются в основании жгутиков и ресничек. По всей вероятности, базальные тельца образуются путем удвоения цен-триолей. Базальные тельца, как и центриоли, являются центрами организации микротрубочек, входящих в состав жгутиков и ресничек.
Жгутики и реснички — органеллы движения у клеток многих видов живых существ. Они представляют собой подвижные цитоплазм этические отростки, служащие либо для передвижения всего организма (многие бактерии, простейшие, ресничные черви) или репродуктивных клеток (сперматозоидов, зооспор), либо для транспорта частиц и жидкостей (например, реснички мерцательных клеток слизистой оболочки носовых полостей и трахеи, яйцеводов и т. д.).
Жгутики эукариотических клеток по всей длине содержат 20 микротрубочек: 9 периферических дуплетов и 2 центральные одиночные. У основания жгутика в цитоплазме располагается базальное тельце.Жгутики имеют длину около 100 мкм и более. Короткие жгутики (10—20 мкм), которых бывает много на одной клетке, называются ресничками.
Скольжение микротрубочек, входящих в состав жгутиков или ресничек, вызывает их биение, что обеспечивает перемещение клетки либо продвижение частиц.
Рибосомы — это мельчайшие сферические гранулы диаметром 15—35 нм, являющиеся местом синтеза белка из аминокислот. Они обнаружены в клетках всех организмов, в том числе прокариотических. В отличие от других органелл цитоплазмы (пластид, митохондрий, клеточного центра и др.) рибосомы представлены в клетке огромным числом: за клеточный цикл их образуется около 10 млн. штук.
В состав рибосом входит множество молекул различных белков и несколько молекул рРНК. Полная работающая рибосома состоит из двух неравных субъединиц (рис. 1.15). Малая субъединица имеет палочковидную форму с несколькими выступами. Большая субь-единица похожа на полусферу с тремя торчащими выступами. При объединении в рибосому малая субъединица ложится одним концом на один из выступов большой субъединицы. В состав малой субъединицы входит одна молекула РНК, в состав большой — три.

Рис. 1.15, Схема строения рибосомы: 1 — малая субъединица; 2 — иРНК; 3 — тРИК; 4 — аминокислота; 5 — большая субьединица; б — мембрана эндоплазматической сети; 7— синтезируемая полипептидная цепь.
В цитоплазме десятки тысяч рибосом расположены свободно (поодиночке или группами) или прикреплены к нитям микротрабекулярной системы, наружной поверхности мембраны ядра и эндоплазматической сети. Они имеются также в митохондриях и хлоропластах.
В процессе синтеза белка рибосома защищает синтезируемый белок от разрушающего действия клеточных ферментов. Механизм защитного действия заключается в том, что часть вновь синтезируемого белка находится в каналоподобной структуре большой субъединицы.
